243

Fig. 6. The two diagrams show the frequency of occurrence of movement (M), search (S),
low tension (LT) and high tension (HT) (vertical axis) during the minute (5-s intervals,
horizontal axis) following the filter-trills (T1 3 5> left) and the arousal-trill (T6, right). Data
are shown for three groups of Guinea fowl on pre- and postoperative sessions. Mean be-
havioural data are shown at intervals for clarity in showing early behavioural changes and
for comparison with preoperative phases (see Figs 4, 5 and text). Prestimulus levels of
behaviour are shown to the left of the vertical axis.
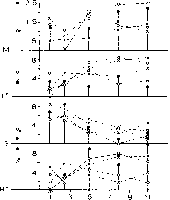
Control group: preoperative not shown, postoperative ∙; anterior group: preoperative
о---o, postoperative о---о; posterior group: preoperativex---x, postoperative x- - -x.
(S)Mixedstimuli.
These stimuli gave rise to stimulus-specific responses. In general the anter-
ior group showed more low tension following any search behaviour than the
posterior group, although both lesion groups showed more low tension than
the sham controls. Specifically, the slow trill (T2) elicited more search in the
lesion groups than in the controls. The arousal trill (T6) elicited much search
behaviour in all groups (Fig. 6). The song (T9) elicited very little search be-
haviour. Instead there was much high tension with the control groups showing
the highest levels (Fig. 7).
Discussion. There are five main points to be made.
Firstly, the test stimuli were recognised as different from the training sti-
muli. This varied by degree with the nature of the stimulus. More search was
observed on the first extinction trial than after the filter-trills. In turn this
level of search was higher than that seen after the ‘novel’ stimuli. (This point
assumes that search and approach represent an activated recognition process
More intriguing information
1. Towards Learning Affective Body Gesture2. Behavior-Based Early Language Development on a Humanoid Robot
3. For Whom is MAI? A theoretical Perspective on Multilateral Agreements on Investments
4. IMPROVING THE UNIVERSITY'S PERFORMANCE IN PUBLIC POLICY EDUCATION
5. The role of statin drugs in combating cardiovascular diseases
6. An Estimated DSGE Model of the Indian Economy.
7. Are Public Investment Efficient in Creating Capital Stocks in Developing Countries?
8. Getting the practical teaching element right: A guide for literacy, numeracy and ESOL teacher educators
9. The name is absent
10. Graphical Data Representation in Bankruptcy Analysis