nd was unambiguous because the boxes were nearly as tall as was the
ubject.
The principal analyses focused on the location of the monkey's first
eacch choice during each of the 50 trials. Search location was coded in one
of two categories (similar to those used by Hermer & Spelke, 1994, 1996).
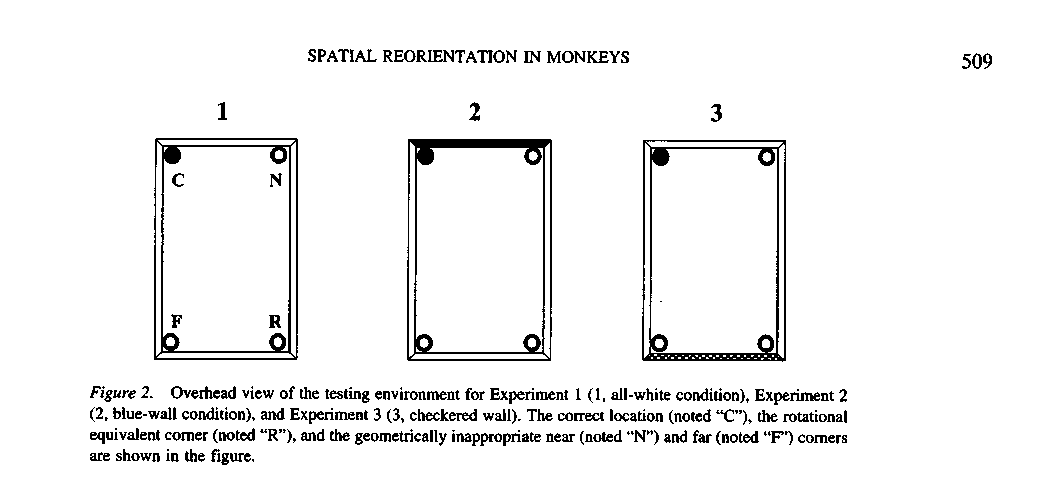
f the subject searched in the correct box (rewarded box, noted "C" for
'correct") or if it was at the rotational equivalent box (at 180° rotation
hrough the center from the rewarded box, noted "R"), this response was
onsidered as a geometrically appropriate choice. Other choices, near the
eward box but geometrically inappropriate or far from the reward box but
pometrically inappropriate (noted respectively, "N" and "F") were called
; eometrically inappropriate choices.
?esults
Table 1 presents the number of first choices performed by the 3
subjects during the 50 trials. For each monkey, these data were
ubjected to a chi-square one-sample test by which we compared
he observed distribution to the theoretical frequency of an equal
listribution in the "geometrically appropriate" and "geometrically
nappropriate" above mentioned categories (i.e., 50% of chance for
:ach one). These results always reached statistical significance,
)rcas, X(1, N = 50) = 20.49; Krill, X2(1, N = 50) = 46.80;
'revet, X2(1, N = 50) = 42.32; p < .001, indicating that the
)bserved distribution between these two categories was not deter-
nined at random. A second chi-square one-sample test was com-
Cable 1
Vumber of Trials (Out of 50) During Experiment 1 for the
subjects as a Function of Their Search Location (Correct,
rotational, Geometrically Inappropriate Corners)
Box
|
Monkey |
CR |
N |
F |
|
)rcas |
21 20 |
4 |
5 |
|
:rill |
25 24 |
1 |
0 |
|
:revet |
23 25 |
2 |
0 |
|
Average (%) |
46 46 |
4.7 |
3.3 |
|
Vote. C = correct; R = |
rotational; N = near misses; |
F = |
far misses. |
IJUIUU uu Liie UUM oouuucu III uic omerveu gevmemcauy appro-
priate category compared with an equal frequency of distributions
of the searches to the two comers of that category (i.e., 50% of
chance for each corner). This analysis indicates that the number of
visits to Corner C was not statistically different from the number
of visits to Corner R, Orcas, X2(1, N = 41) = 0.02; Krill, X2(1, N
= 49) = 0.02; Crevet, X2(1, N= 48) = 0.08; p > .05. This result
shows that the correct and the geometrically equivalent corners
were chosen with no specific preference.
Data analyses of the first 10 trials for each subject are reported
in Table 2 (Experiment 1). These data were submitted to a logistic
regression to test whether the scores for the 3 subjects, during
the 10 first trials, show differences with the results of the entire
experimental session (i.e., 50 trials). The analyses show that the
performance of each subject is not statistically different within the
same experiment, Orcas, x(1, N = 10) = 0.07; Krill, X2(1, N =
10) = 0.01; Crevet, X2(l, N = 10) = 0.25; p > .05. Thus, we can
conclude that no improvement of the monkeys' performance takes
place across trials.
Discussion
Results of Experiment 1 show that the monkeys rely on the
geometry of the room to reorient. Indeed, in the present homoge-
neous environment (white-room condition), monkeys confined
their search almost exclusively to both geometrically correct cor-
ners (the correct corner and the corner of the opposite side of the
chamber), searching in these two locations with an equal fre-
quency. Thus, like mature rats, toddlers, and adults, rhesus mon-
keys are able to reorient by reference to the shape of the environ-
ment (Cheng, 1986; Hermer & Spelke, 1996).
Our results also indicate that the disorientation procedure used
in this experiment is efficient and that no other landmark but the
shape of the room is used by the subject to reorient in the exper-
imental environment. Neither visual information within the exper-
imental room nor nonvisual information from outside the room
served as cues for reorientation. If any external or internal cue had
been used, then monkeys would certainly have been able to locate
the target without ambiguity.
More intriguing information
1. Feature type effects in semantic memory: An event related potentials study2. The Role of Land Retirement Programs for Management of Water Resources
3. The name is absent
4. Momentum in Australian Stock Returns: An Update
5. Program Semantics and Classical Logic
6. The name is absent
7. Yield curve analysis
8. Quelles politiques de développement durable au Mali et à Madagascar ?
9. The name is absent
10. PROPOSED IMMIGRATION POLICY REFORM & FARM LABOR MARKET OUTCOMES