and smaller scales, a process Wallace and Wallace (2008) char-
acterize as mesoscale resonance, a generalization of the Bald-
win effect. See that reference for details, broadly based on
the Tuning Theorem.
14.3 Ecosystems farm organismal develop-
ment
The environmental and ecosystem farming of development
may not always be benign.
Suppose we can operationalize and quantify degrees of both
overfocus/inattentional blindness (IAB) and of overall struc-
ture/environment distortion (D) in the actions of a highly
parallel cognitive epigenetic regulatory system. The essential
assumption is that the (internal) dual information source of a
cognitive structure that has low levels of both IAB overfocus
and structure/environment distortion will tend to be richer
than that of one having greater levels. This is shown in figure
1a, where H is the source uncertainty dual to internal cogni-
tive process, X = IAB , and Y = D. Regions of low X, Y ,
near the origin, have greater source uncertainty than those
nearby, so H(X, Y ) shows a (relatively gentle) peak at the
origin, taken here as simply the product of two error func-
tions.
We are, then, particularly interested in the internal cogni-
tive capacity of the structure itself, as paramatized by degree
of overfocus and by the (large scale) distortion between im-
plementation and impact. That capacity, a purely internal
quantity, need not be convex in the parameter D, which is
taken to characterize interaction with an external environ-
ment, and thus becomes a context for internal measures.
The generalized Onsager argument, based on the homology
between information source uncertainty and free energy, as
explained more fully in the Appendix, is shown in figure 1b.
S = H(X,Y) - XdH/dX - Y dH/dY, the ‘disorder’ analog
to entropy in a physical system, is graphed on the Z axis
against the X - Y plane, assuming a gentle peak in H at the
origin. Peaks in S , according to theory, constitute repulsive
system barriers, which must be overcome by external forces.
In figure 1b there are three quasi-stable topological resilience
modes, in the sense of Wallace (2008b), marked as A, B, and
C . The A region is locked in to low levels of both overfocus
and distortion, as it sits in a pocket. Forcing the system
in either direction, that is, increasing either IAB or D, will,
initially, be met by homeostatic attempts to return to the
resilience state A, according to this model.
If overall distortion becomes severe in spite of homeostatic
developmental mechanisms, the system will then jump to
the quasi-stable state B , a second pocket. According to the
model, however, once that transition takes place, there will
be a tendency for the system to remain in a condition of high
distortion. That is, the system will become locked-in to a
structure with high distortion in the match between struc-
ture implementation and structure impact, but one having
lower overall cognitive capacity, i.e., a lower value of H in
figure 1a.
The third pocket, marked C , is a broad plain in which both
S=H-XdH∕dX-YdS∕dY
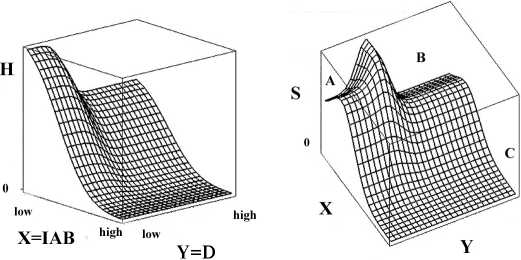
Figure 1: a. Source uncertainty, H, of the dual information
source of epigenetic cognition, as parametized by degrees of
focus, X = IAB and distortion, Y = D, between implemen-
tation and actual impact. Note the relatively gentle peak at
low values of X, Y . Here H is generated as the product of two
error functions. b. Generalized Onsager treatment of figure
1a. S = H(X, Y)-XdH/dX-Y dH/dY. The regions marked
A, B , and C represent realms of resilient quasi-stability, di-
vided by barriers defined by the relative peaks in S . Transi-
tion among them requires a forcing mechanism. From another
perspective, limiting energy or other resources, or imposing
stress from the outside - driving down H in figure 1a, would
force the system into the lower plain of C , in which the sys-
tem would then become trapped in states having high levels
of distortion and inattentional blindness/overfocus.
15
More intriguing information
1. The name is absent2. Locke's theory of perception
3. The name is absent
4. Tax Increment Financing for Optimal Open Space Preservation: an Economic Inquiry
5. The name is absent
6. The name is absent
7. The name is absent
8. The quick and the dead: when reaction beats intention
9. The name is absent
10. AJAE Appendix: Willingness to Pay Versus Expected Consumption Value in Vickrey Auctions for New Experience Goods