composed of several segregated channels, each of
which being involved in a particular functional loop
with the frontal cortex and its corresponding part of
the thalamus to achieve selection of action. Basically,
a thalamo-cortical network (bidirectional excitatory
connections) is tonically inhibited by the BG output
(here the internal segment of the Globus Pallidus
GPi). The core mechanism of BG is disinhibition,
which means that inhibition of the GPi (by the Stria-
tum or by the external part of the Globus Pallidus
GPe) disinhibits the corresponding part of the tha-
lamus, thus allowing reciprocal excitation between
the thalamus and the cortex. This mechanism has
already been used in the attention-switching system.
Without giving too much detail, there are two op-
posite pathways in the BG architecture: the direct
pathway that favorizes disinhibition and an indirect
one which prenvents disinhibition. The balance be-
tween these two pathways is ensured by the dopamin-
ergic signal produced by the Substancia Nigra pars
compacta (SNc).
This complex architecture with bidi-
rectional connections, internal loops and
dopaminergic modulation allows to learn
sequences and do selection of action
(Berns and Sejnowski, 1998, Gurney et al., 2004).
We will describe here how our model is able to learn
simple sequences like ordinal numbers (cf. Figure
2).
⅛ ⅛
“zero” “one”

“two” “three”
Figure 3: The distributed representation of the phoneti-
cal words ’zero’ ’one’ ’two’ ’three’ in the number map.
- We first organized a neural map called number
representation map with phonological inputs repre-
senting numbers (zero, one, two, three). The hearing
of one of these numbers therefore implies the appear-
ance of a bubble of activity in the number map at
a given location on the map (Figure 3). Please note
that this coding is not compact and would not scale
to large numerosities.
- We then present successively the four numbers
to the BG model with a phasic burst of dopamine in
SNc at the time of the switch. As dopamine stands
for a kind of “reward” signal (Schultz et al., 1992),
we can justify this by saying that hearing a voice
(one’s mother’s voice for example) is intrinsically re-
warding.
- The inner dynamics of the system (not described
here) ensure that the association between the cortical
representation of a number and its follower is learned
by the connections between stn and gpe.
After learning, when some stn neurons are active
for the current number (via their cortical inputs),
they tend to excite gpe at the location of the next
number, which in turn inhibits gpi. This artificially
creates disinhibition in the thalamus that can be used
to predict the next number.
- A high tonic level of dopamine favorizes the di-
rect pathway so that the representation of the cur-
rent number in the number map is stable even with-
out any phonological input (it is mainly the same
reverbatory mechanism as in the attention-switching
system).
- A sudden depletion of dopamine leads to an ad-
vantage for the indirect pathway that predicts the
location of the next number: the cortical representa-
tion in the number map switches to the next number
without any corresponding phonological input, like a
kind of mental voice.
5. Merging the two Systems
1
0.9
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0
"reward_trace"

b)
a)
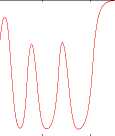
Figure 4: a) Timecourse of the reward signal when three
objects are presented in the visual scene. Each depletion
corresponds to a switch of focus of attention. b) Corre-
sponding image.
We briefly showed how the sequence-learning sys-
tem could learn to reproduce a phonological sequence
of numbers. After a tonic level of dopamine is applied
in SNc to start the counting task, each dopamine
depletion switches the cortical representation to the
following number. If we add a inhibitory connec-
tion from the focus map in the attention-switching
mechanism to its reward unit, the system focuses
each salient point in the image once and then stops.
The timecourse of the activity of the reward unit
is shown in Figure 4 for three ob jects. It is al-
most the same timecourse needed for the SNc of
the sequence-learning mechanism to reproduce the
learned sequence.
Having noticed this analogy, our idea was to link
the two systems only by their dopaminergic unit: the
reward unit of the attention-switching system be-
comes the SNc unit of the other model and then con-
trols the restitution of the learned sequence. In other
words, each time an ob ject is focused, the current
number is incremented. At the end, when no more
salient object can be found, the sequence-learning
127
More intriguing information
1. The constitution and evolution of the stars2. Modelling the Effects of Public Support to Small Firms in the UK - Paradise Gained?
3. Credit Market Competition and Capital Regulation
4. A NEW PERSPECTIVE ON UNDERINVESTMENT IN AGRICULTURAL R&D
5. The Global Dimension to Fiscal Sustainability
6. The name is absent
7. Biological Control of Giant Reed (Arundo donax): Economic Aspects
8. Categorial Grammar and Discourse
9. The Making of Cultural Policy: A European Perspective
10. Industrial Cores and Peripheries in Brazil