434
(Oades et al. 1992). Increased DA and NA activ-
ity may reflect that their response to medication
had not entirely stabilized. Independent of the bio-
chemical dimension it appear s that acutely ill schi-
zophrenics can experience problems in determin-
ing stimulus-relevance, as has also been reported
with the related latent inhibition task (Baruch et
al. 1988).
Signal processing and Hionoaminergic
modulation
I have just implied a DA role in schizophrenic prob-
lems with attention. From the problems of schizo-
phrenics with thought disorder and attention
(reviews: Oades 1982; Straube and Oades 1992)
and from basic considerations (Fig. 3) it may be
assumed that a number of transmitter systems are
involved in mediating the several components of
an attentional system. Selective processing is
bound to involve excitatory and inhibitory amino
acids in relatively local cortical mediation, Ionger-
Ioop subcortical comparison and to be modulated
by long-axon ascending monamines. Let us con-
sider the monoamines first.
Serotonin (5HT) can exert a form of volume
control. Deliver a pulse burst to the raphe in the
cat and there is a facilitation of the potentials one
can evoke in the hippocampal perforantpath 5-140
msec later. This effect depends on the state of wake-
fulness (Trulson and Jacobs 1979; Srebro et al.
1982). The interval is one relevant to the elicitation
of LTP and approximates the theta frquency. It is
possible to predict learning speed in rabbits from
small changes of theta (Thompson et al. 1980).
Small decreases of theta frequency follow toxic
raphe lesions and increases follow raphe stimula-
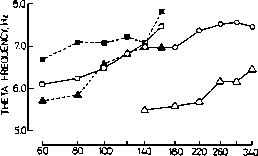
tion (e.g. Graeff et al. 1980, Fig. 5). This illus-
trates one aspect of the potential involvement of
5HT in selective processing, which as mentioned
above may be reflected in the Nl-P2 amplitude
and changes of this seen in psychiatric patients.
Noradrenaline (NA) function is often described
in terms of tuning. There must be at least two in-
puts to a given brain region vying for control of
the output. NA biases the relative control of activ-
ity in the region and thus tunes the response (re-
view, Oades 1985). The classic report leading to
this formulation came from Segal and Bloom
(1976). Essentially they found that suppression of
firing of hippocampal cells in the rat by a tone
would habituate, but could be reinstated through
electrical stimulation of the locus Coeruleus, ori-
gin of the NA innervation. However if the tone
acted as a conditioned stimulus, then pairing tone
with Coeruleus stimulation increased hippocampal
firing.
STIMULUS INTENSITY, μA
TRIAL OF 5th CR
Fig. 5. Left: Increasing frequencies Ofhippocampal theta rhyhtm produced by increasing intensities of electrical stimulation of
the median raphe (50 Hz, 0.5 msec, 5 sec train). Each point is the mean of two tests in one session or an individual rat. (From
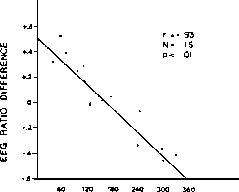
Graeff et al. 1980 with permission of the author and Pergamon Press.) Right: Scatterplot and regression line showing a signifi-
cant relation between changes in the ratio of high to low EEG and the number of training trials required to attain the 5th
conditioned response. Pre-task EEG samples showed the more theta, the faster the learning raste r = +0.72: task, — conditioning
nictitating membrane response to air puffs in the rabbit. (From Thompson et al. 1980 with permission of the author and
publisher.)
More intriguing information
1. DISCRIMINATORY APPROACH TO AUDITORY STIMULI IN GUINEA FOWL (NUMIDA MELEAGRIS) AFTER HYPERSTRIATAL∕HIPPOCAMP- AL BRAIN DAMAGE2. Sustainability of economic development and governance patterns in water management - an overview on the reorganisation of public utilities in Campania, Italy, under EU Framework Directive in the field of water policy (2000/60/CE)
3. Public infrastructure capital, scale economies and returns to variety
4. Moi individuel et moi cosmique Dans la pensee de Romain Rolland
5. Imperfect competition and congestion in the City
6. The name is absent
7. The name is absent
8. he Effect of Phosphorylation on the Electron Capture Dissociation of Peptide Ions
9. Voting by Committees under Constraints
10. The name is absent