36
(59.9%) and 0P2 (59.8%) were significantly active, with a smaller fraction of active
voxels in OP3 (19.1%) and OP4 (21.7%).
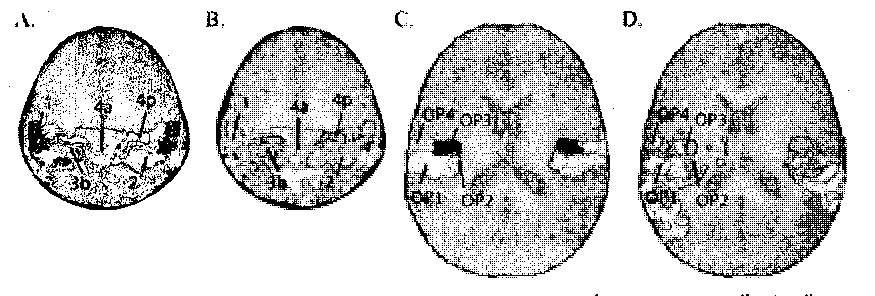
Figure 7. Relationship between Cytoarchitectonically defined areas and somatosensory Vibrotactile
activation. A, Cytoarchitectonic regions in and near Sl from the Anatomy Toolbox (z=54 mm)
(Eickhoff, et al. 2005; Geyer, et al. 1999; Grefkes, et al. 2001). B, Average somatosensory activation
(orange-to-yellow color scale) under cytoarchitectonie boundaries (colored outlines). C,
Cytoarchitectonic regions in and near S2 (z=18 mm) (Eickhoff, et al. 2006a; Eickhoff, et al. 2006b). O,
Average somatosensory activations under cytoarchitectonie boundaries.
Discussion
These experiments demonstrate that simple Vibrotactile stimuli evoke robust
BOLD fMRI responses in MST but not in MT. A potential source for Vibrotactile responses
in MST is a projection from the ventral intraparietal area (VIP). In nonhuman primates,
VlP receives input from hand and arm regions of S2, and in turn
VIP projects to MST (Lewis and Van Essen, 2000). Connections from macaque VIP to MT
are much sparser than those from VIP to MST, perhaps reflecting the weaker
Vibrotactile responses in MT than MST observed in the present study. Although the
connectivity of human MST is uncertain, studies using diffusion tensor
fractography (Rushworth, et al. 2006) and functional effective connectivity (Peltier, et al.
2007) offer promising avenues of exploration. Additional support for VIP contributions
More intriguing information
1. The migration of unskilled youth: Is there any wage gain?2. The name is absent
3. Putting Globalization and Concentration in the Agri-food Sector into Context
4. Clinical Teaching and OSCE in Pediatrics
5. Examining Variations of Prominent Features in Genre Classification
6. ARE VOLATILITY EXPECTATIONS CHARACTERIZED BY REGIME SHIFTS? EVIDENCE FROM IMPLIED VOLATILITY INDICES
7. The name is absent
8. Estimating the Technology of Cognitive and Noncognitive Skill Formation
9. Using Surveys Effectively: What are Impact Surveys?
10. Models of Cognition: Neurological possibility does not indicate neurological plausibility.