Rate at which monkeys sample the sensory environment
347
What figure 3 shows is that the slopes of the survivorship curves—and thus the
half-lives of the distributions—change as a function of colour and noise. In
experiment 1 the approximate half-lives of the mean distributions were 5 ∙ 5 s in red
light and 7∙9 s in blue light; in experiment 2 they were 5-2 s in 90 dB noise and
7∙8 s in 60 dB noise. All seven individual monkeys reacted to colour and noise in
the same way (half-life in red < half-life in blue; half-life in 90 dB < half-life in
60 dB), giving a significance to each result of p < 0∙01.
4 Discussion
These results could be ‘explained’, in the same way that we explained the results of
our last series of experiments (Humphrey and Keeble 1977), in terms of an influence
of the sensory environment on the passage of subjective time. Thus it might be
suggested that monkeys have a subjective clock which not only runs faster in red
light than in blue but also—on the evidence of the present study—runs faster in
90 dB noise than in 60 dB noise. The trouble with this explanation is that it does
not really get to grips with the behavioural phenomenon: it explains the effects of
colour and noise on a pattern of behaviour which itself remains unexplained. Just
what were the monkeys about when they moved back and forth between the
chambers?
No one who observed the monkeys could have thought that they were merely
indulging in aimless motor activity. Each of their moves looked purposeful. Yet,
since the two chambers were identical, no obvious purpose was in fact achieved by
moving. Can the monkeys have gone on shuttling to and fro in the vain expectation
that next time there might be something different about the other chamber?
Look at the Necker Cube in figure 4. You see it first this way round, then that
way, then this way again. Your mind is never satisfied that it has reached the right
interpretation. No matter that you have already seen both aspects of the cube a
dozen times, you go on vainly ‘sampling’ each of the possible alternatives. So,
perhaps, with the monkeys in the testing apparatus: able to be in only one place at
one time, they could never be sure that they were not missing something in the
other chamber—and every so often the urge came over them to check on it.
In the real world such periodic checks would have a clear function. The real
world, unlike the testing apparatus, cannot be counted on to remain stable over time.
Things which are out of sight are out of ken and if a monkey is to keep itself
informed it must continually sample hidden bits of its environment. Such sampling
may sometimes amount to no more than a glance over the shoulder, sometimes a
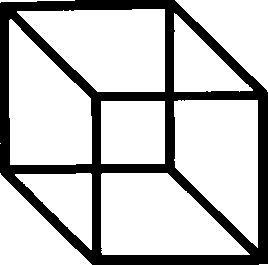
Figure 4. Necker Cube. Note that a recent attempt to characterise the probability distribution of
temporal fluctuations in the perception of this figure concludes that “the underlying process may
be described by a simple Poisson model” (De Marco et al 1977).
More intriguing information
1. DURABLE CONSUMPTION AS A STATUS GOOD: A STUDY OF NEOCLASSICAL CASES2. Picture recognition in animals and humans
3. Connectionism, Analogicity and Mental Content
4. Innovation Trajectories in Honduras’ Coffee Value Chain. Public and Private Influence on the Use of New Knowledge and Technology among Coffee Growers
5. Meat Slaughter and Processing Plants’ Traceability Levels Evidence From Iowa
6. Ventas callejeras y espacio público: efectos sobre el comercio de Bogotá
7. Developmental Robots - A New Paradigm
8. The Dictator and the Parties A Study on Policy Co-operation in Mineral Economies
9. The name is absent
10. Private tutoring at transition points in the English education system: its nature, extent and purpose