Activation of s28-dependent transcription by CRP 1107
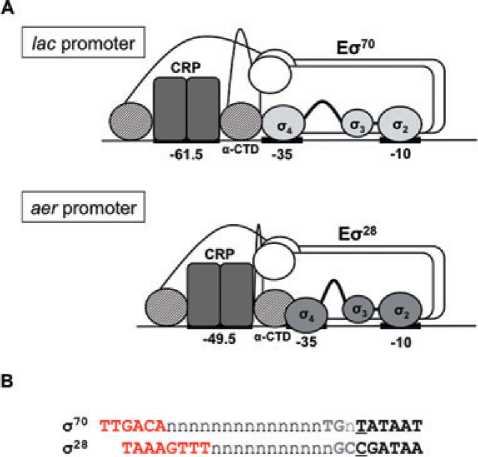
Fig. 8. Organization of the CRP-RNA polymerase-DNA complex
at the aer promoter.
A. Comparison of the CRP-RNA polymerase-DNA complexes at
s70- and s28-dependent promoters. The upper panel shows a
schematic diagram of the CRP-Es70-DNA ternary complex at the
Class I CRP-activated lac promoter, where CRP binds at a site
centred 61.5 bp upstream of the transcript start site. The lower
panel shows a model for the organization of the CRP-Es28-DNA
ternary complex at the aer promoter, where CRP binds at a site
centred 49.5 bp upstream from the transcript start site. Domain 4 of
s28 is proposed to contact the DNA further downstream at the aer
promoter than does domain 4 of s70 at the lac promoter, and this
relieves a potential clash with downstream-bound aCTD.
B. Spacing between the -10 and -35 elements at s70- and
s28-dependent promoters. The figure shows the s70 and s28
promoter consensus sequences, aligned by the position of the
upstream edge of the core -10 element (underlined). The core -10
element is shown in black, the extended -10 element in grey and
the -35 element in red. Each base in the spacer region is
represented by an ‘n’.
model, illustrated in Fig. 8A, suggests that domain 4 of s28
is offset by rotation and translocation around the surface
of the promoter DNA. It is quite possible that this is its
‘normal’ position at s28-dependent promoters, but this will
require further experimental evidence.
Although E. coli contains hundreds of transcription acti-
vators, there are few examples of factor-dependent acti-
vation of promoters recognized by alternative s factors
such as s28. Transcription from promoters served by alter-
native s factors is mostly regulated by controlling the
expression and activity of the s factor itself, and by the
very stringent promoter recognition properties of the alter-
native s factors. It is generally accepted that control over
most flagellar genes is exerted by regulating the expres-
sion and activity of FlhDC, s28, and the anti-s factor, FlgM
(Chilcott and Hughes, 2000; Keseler et al., 2009). Our
findings show that transcription activators can also play
an important role in controlling transcription by Es28.
The similar organization of the aer and trg promoters
suggests a common mechanism of direct activation by
CRP. However, we found no evidence for direct regulation
by CRP at the six other well-characterized s28-dependent
promoters in E. coli K-12. This raises the question of why
CRP should directly regulate transcription of aer and trg,
particularly when CRP indirectly regulates transcription of
all genes in the flagellar cascade by controlling expres-
sion of the master regulator, FlhDC. Expression of FlhDC
is tightly regulated by multiple transcription factors, includ-
ing CRP, H-NS and OmpR, and hence the genes of the
flagellar cascade are regulated in response to many dif-
ferent environmental inputs (Shin and Park, 1995; Sout-
ourina et al., 1999). It is possible that expression of aer
and trg is required only in response to conditions that
induce CRP activity and not in response to other signals
that induce the flagellar cascade. Alternatively, expression
of aer and trg may need to be upregulated to a greater
extent than other genes when CRP activity is induced.
This may be particularly important when levels of Es28 are
low. In these conditions, recruitment of Es28 by CRP may
ensure that the aer and trg transcription units are prefer-
entially expressed compared with other s28-dependent
genes. Note that aer and trg encode homologous methyl-
accepting chemotaxis regulator-type proteins, which inter-
act with the flagellar motor via the CheA/CheY signalling
pathway to control the direction of bacterial movement in
response to different chemical or energetic signals (Taylor
et al., 1999). The direct involvement of CRP in their regu-
lation may therefore simply be a reflection of the role of
CRP in facilitating the acquisition and metabolism of nutri-
ents other than glucose (Kolb et al., 1993).
Experimental procedures
Strains, plasmids and promoter constructs
The E. coli K-12 strains, plasmids and promoter fragments
used in this study are listed in Table 2. Standard recombinant
DNA techniques were used throughout and all the oligonucle-
otide primers used are listed in Table S1.
The DfliA derivatives of strains M182 and M182 Dcrp were
constructed by P1 transduction of a fliA::kan mutation from
strain JW1907-1. The kanamycin resistance marker was sub-
sequently removed by expressing the FLP recombinase from
plasmid pCP20, as described by Cherepanov and Wacker-
nagel (1995), and the presence of the deletion was confirmed
by colony PCR using primers D56550 and D56551.
Plasmid pKXH100 was constructed by cloning an NdeI-
XhoI fragment carrying the fliA coding sequence from E. coli
K-12 strain MG1655, amplified by PCR using primers
D57845 and D57846, into plasmid pET21a (Novagen). As a
result of leaky expression, genes cloned under the control of
the T7 promoter in pET21a are expressed even in strains that
do not produce T7 RNA polymerase, including M182 (Wu
et al., 2005). This activity is independent of the presence of
the inducer IPTG (K. Hollands, unpubl. results).
© 2009 The Authors
Journal compilation ©2009 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1098-1111
More intriguing information
1. Public Debt Management in Brazil2. The name is absent
3. BILL 187 - THE AGRICULTURAL EMPLOYEES PROTECTION ACT: A SPECIAL REPORT
4. The name is absent
5. The Shepherd Sinfonia
6. How do investors' expectations drive asset prices?
7. The Integration Order of Vector Autoregressive Processes
8. Temporary Work in Turbulent Times: The Swedish Experience
9. Artificial neural networks as models of stimulus control*
10. Developments and Development Directions of Electronic Trade Platforms in US and European Agri-Food Markets: Impact on Sector Organization