Activation of s28-dependent transcription by CRP 1105
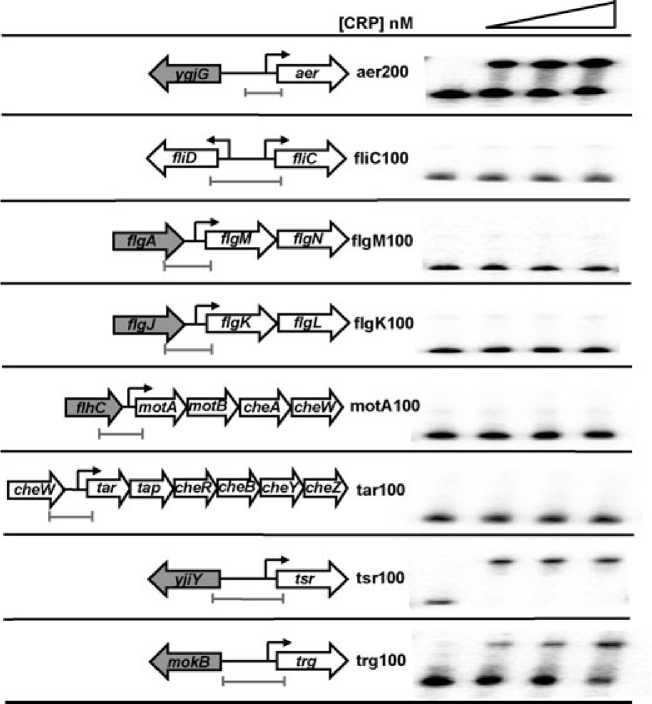
Fig. 6. Binding of CRP to s28-dependent
promoters in vitro. The left hand panels show
schematic diagrams of the regulatory regions
of the eight well characterized s28-dependent
promoters from E. coli K-12 (not to scale).
s28-dependent genes are indicated by open
arrows, while genes that are not known to be
s28-dependent are shown as grey arrows.
Black lines denote intergenic regions, and
black arrows show the locations of known or
putative s28-dependent promoters. Grey bars
indicate the extent of the EcoRI-HindIII
promoter fragments used in this work.
The right hand panels show the results of
electromobility shift assays using the
end-labelled promoter fragments, incubated
with 0, 50, 100 or 200 nM CRP, in the
presence of 0.2 mM cAMP.
latory regions revealed that the spacing between the pre-
dicted DNA sites for CRP and the different elements of the
two s28-dependent promoters is identical (Fig. 7A).
To measure the effect of CRP on expression from the
trg regulatory region, the trg100 promoter fragment was
cloned into pRW50, and the activity of each promoter
::IacZfusion was measured in the CRP+ FliA+, CRP- FliA+
and CRP+ FliA- backgrounds. Recall that, in the condi-
tions used in our experiments, direct effects of CRP on
transcription of promoter::lacZ fusions in pRW50 can be
measured independent of the effect of CRP on s28 levels.
Results illustrated in Fig. 7B indicate that expression from
the trg regulatory region, like the aer promoter, is depen-
dent on s28 and is activated by CRP. The conservation of
the spacing between the DNA site for CRP and the -10
and -35 elements at the aer and trg promoters suggests
that the mechanisms of transcription activation at the two
promoters are similar. Interestingly, the trg and aer genes
encode homologous proteins with similar functions. While
Aer is an energy sensor that controls responses to redox
signals, Trg is a chemosensor that responds to the
monosaccharides ribose and galactose (Taylor et al.,
1999).
Conclusions
Here we have described the first examples of direct acti-
vation by CRP of promoters served by RNA polymerase
holoenzyme containing the flagellar sigma factor, s28. We
showed that transcription of the E. coli K-12 aer gene is
driven by a single s28-dependent promoter, which is acti-
vated by CRP binding to a single site positioned 49.5 base
pairs upstream of the transcript start site. This location
appears optimal for activation. This is in contrast to the
situation at previously studied Class I and Class II CRP-
dependent promoters where the optimal locations for acti-
vation by CRP are positions -61.5 and -41.5 respectively,
and where CRP activates only very weakly when bound to
a site centred near position -50 (Gaston et al., 1990). Our
results argue that the spacing requirements for CRP-
dependent activation at promoters served by Es28 differ
from those at promoters served by Es70. It is possible that
promoters recognized by some other alternative s factors
also require CRP binding at unusual locations. For
example, at the s38-dependent csiD promoter, CRP acti-
vates optimally from a DNA site centred at position -68.5
(Germer et al., 2001). From this position, or a site located
© 2009 The Authors
Journal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1098-1111
More intriguing information
1. Macroeconomic Interdependence in a Two-Country DSGE Model under Diverging Interest-Rate Rules2. Competition In or For the Field: Which is Better
3. Testing Hypotheses in an I(2) Model with Applications to the Persistent Long Swings in the Dmk/$ Rate
4. DISCRIMINATORY APPROACH TO AUDITORY STIMULI IN GUINEA FOWL (NUMIDA MELEAGRIS) AFTER HYPERSTRIATAL∕HIPPOCAMP- AL BRAIN DAMAGE
5. XML PUBLISHING SOLUTIONS FOR A COMPANY
6. Fiscal Sustainability Across Government Tiers
7. CROSS-COMMODITY PERSPECTIVE ON CONTRACTING: EVIDENCE FROM MISSISSIPPI
8. A Dynamic Model of Conflict and Cooperation
9. The name is absent
10. Skills, Partnerships and Tenancy in Sri Lankan Rice Farms