11b-HSD1 is essential for human adipogenesis . I J BUJALSKA and others 303
І 400
I 300
S
S 200
CD
§ 100
о
p 0
о
03PD
***

DayO Day3 Day 5 Day 7 Day 9 Day 16
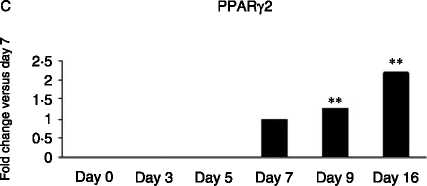
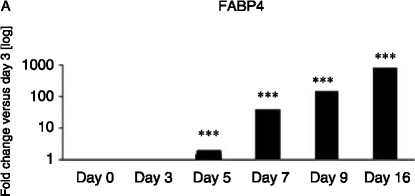
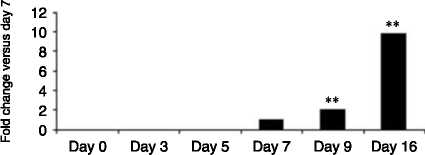
Figure 5 Gene expression analysed by real-time PCR of adipogenic markers; (A) FABP4: (log scale) 2-fold increase on day 5,
38-fold on day 7, 142-fold on day 9 and 870-fold on day 16 versus day 3; (B) G3PD: 4.5-fold on day 7, 22-fold on day 9 and
380-fold on day 16 versus day 5; (C) PPARg2: 1.3-fold on day 9 and 2.2-fold on day 16 versus day 7; and (D) GLUT-4: 2.1-fold
on day 9 and 9.8-fold on day 16 versus day 7 across Chub-S7 cell differentiation. P values: **P!0.01, ***P!0.001.
GLUT4
D
defined the permissive role of GCs in inducing adipocyte
differentiation. Here, we extend those observations to a novel
transformed human adipocyte cell line that should greatly
enhance in vitro human-based adipocyte research studies. Within
5 days of incubation in chemically defined media comprising
insulin, PPARg agonist and glucocorticoid, impressive differ-
entiation was observed in Chub-S7 cells as assessed by markers
including FABP4, G3PD and adipocyte-specific genes such as
GLUT-4 and PPARg2. Adipogenesis was an ongoing process
up to 16 days in culture with intracellular lipid stores confirmed
by oil red O staining. No significant changes were observed in
GRa expression during this differentiation phase; earlier studies
had reported increased GR expression in omental versus
subcutaneous adipose tissue and had argued that this may be
the one factor explaining the predilection of glucocorticoids for
visceral obesity (Bronnegard et al. 1990). However, more
recently our array and real-time PCR analyses failed to
demonstrate any difference in GR expression between human
omental and subcutaneous preadipocytes (Bujalska et al. 2006)
and adipose tissue (unpublished data). Together with the data in
this study, it seems unlikely that changes in GR expression per se
are important in the adipogenesis process.
At a pre-receptor level our group has focussed on the role of
11b-HSD1 and the regeneration of cortisol from inactive
cortisone in human adipose tissue. Previously we have
demonstrated increased expression of 11b-HSD1 in omental
compared with subcutaneous depots (Bujalska et al. 1997b), and
importantly a switch in the directionality of enzyme activity
from predominant dehydrogenase to oxo-reductase (Bujalska
et al. 2002a), probably as a result of induction of the NADPH
donor source for 11b-HSD1 within the endoplasmic reticulum
(ER) lumen, H6PDH (Ozols 1993). In support of these
observations, in this study, the rise in H6PDH mRNA levels
probably explained the increase in 11b-HSD1-mediated oxo-
reductase activity between days 3 and 9 in addition to an increase
in 11b-HSD1 mRNA levels. Furthermore, decreased H6PDH
mRNA expression seen on day 16 might account for the decline
in 11b-HSD1 oxo-reductase activity despite high mRNA. The
impressive increase in 11b-HSD1 expression across adipocyte
differentiation has been noted recently in mouse 3T3-L1 cells
(Kim et al. 2007), as it has been done in other differentiating cell
systems including osteoblasts (Eijken et al. 2005). In both cases
cells have been incubated with glucocorticoids that themselves
positively regulate 11b-HSD1 (Bujalska et al. 1997b); it is not
possible from these or previous data sets to conclude whether or
not the increased 11b-HSD1 expression is glucocorticoid
mediated or a manifestation of the differentiation process per se.
Nevertheless, 11b-HSD1 expression, resulting in a function-
ally active oxo-reductase, was clearly linked to the differentiation
phenotype. Incubation of Chub-S7 cells with inactive steroid,
cortisone, induced a degree of adipocyte differentiation similar
to that observed with cortisol. This could be explained on the
basis of 11b-HSD1-mediated oxo-reductase activity with a Km
for cortisone in Chubb-S7 cells of 100 nM which is similar to
the reports in other 11b-HSD1-expressing cell systems
(Monder & Lakshmi 1989, Ricketts et al. 1998) including
studies on the recombinant enzyme (KmZ333 nM; Walker et al.
2001, Shafqat et al. 2003). Evaluation of the competitive
11b-HSD1 inhibitor, PF-877423, indicated selectivity for
the type 1 11b-HSD isoform with a Ki£0.2 nM in the
recombinant assay and IC50w5 nM in Chubb-S7 cells.
Selectivity of the inhibitor towards 11b-HSD1 isoform but not
11b-HSD2 is crucial as patients with impairment in the latter
are presented with severe hypertension, (White et al. 1997).
www.endocrinology-journals.org
Journal of Endocrinology (2008) 197, 297-307
More intriguing information
1. Flatliners: Ideology and Rational Learning in the Diffusion of the Flat Tax2. Rent-Seeking in Noxious Weed Regulations: Evidence from US States
3. Evaluating the Impact of Health Programmes
4. The Environmental Kuznets Curve Under a New framework: Role of Social Capital in Water Pollution
5. Structural Breakpoints in Volatility in International Markets
6. Perfect Regular Equilibrium
7. Connectionism, Analogicity and Mental Content
8. The name is absent
9. The name is absent
10. Conservation Payments, Liquidity Constraints and Off-Farm Labor: Impact of the Grain for Green Program on Rural Households in China